FectoPRO®
FectoPRO is a transfection reagent specifically optimized for transient protein and antibody expression in CHO and HEK293 cells.
![]()
![]()
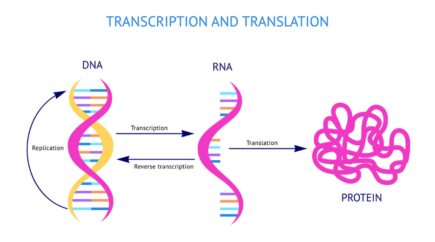
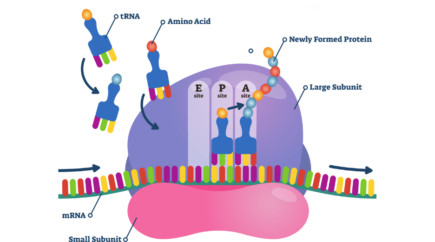
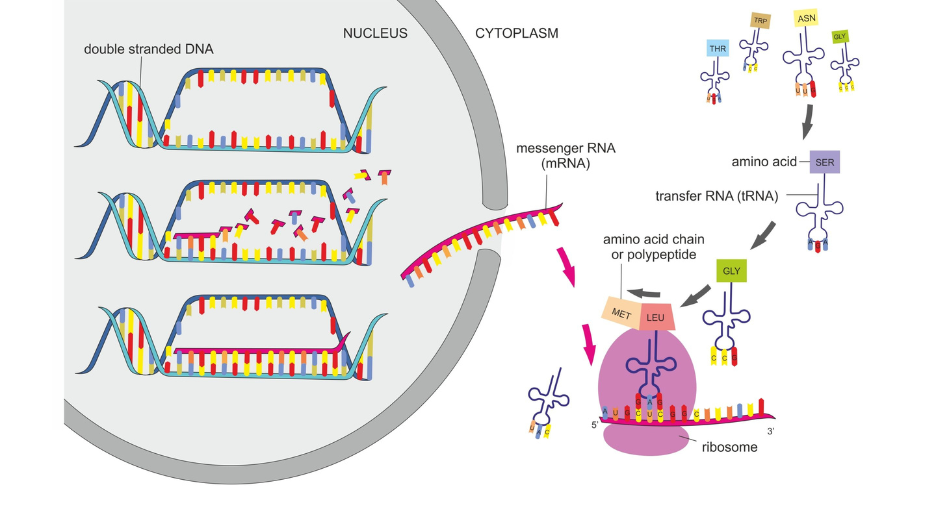
Involvement of three proteins known as initiation factors IF1, IF2 and IF3 completes the initiation complex. The conformation of this multimolecular complex brings the AUG start codon of the mRNA in front of the P site of the 30S subunit. IF2 is then recruiting the initiator tRNA to the complex, which in turn allows the recruitment of the large subunit (50S) to form the complete and active 70S ribosome3.4. In bacteria the initiator tRNA is carrying a N-formylmethionine that will be the first amino acid in the nascent peptide and will be removed during the elongation step.

FectoPRO is a transfection reagent specifically optimized for transient protein and antibody expression in CHO and HEK293 cells.
jetOPTIMUS® is an innovative cationic nanotechnology developed to improve DNA transfection efficiency in hard-to-transfect cells.
jetPRIME® is a versatile DNA and siRNA transfection reagent that ensures high DNA transfection efficiency and excellent gene silencing.

Polyplus provides an integrated service for design and production of tailor-made DNA plasmid through our unique and Innovative assembly technology.

Polyplus offers a comprehensive range of plasmid manufacturing services from DNA plasmid design to plasmid DNA manufacturing and analytical testing.